


Responses of Photosystem II of White Elm to UV-B Radiation Monitored by OJIP Fluorescence Transients
2011-09-22
White elm (Ulmus pumila L.) is widely found in the semi-arid and arid regions of China. It is an important hardwood with high economic value. Its fruit is a dislike samara, being composed of a compressed nutlet (seed) surrounded by a membranous wing. The seed is centrally embedded in the wings, so the photosynthetic performance in samara wings is closely related with the quality of the seed inside.
In the semi-arid and arid regions of China, the UV-B (280-315 nm) radiation reaching the earth’s surface is kept at high levels for most time. What is more important, the solar UV-B radiation (UVBR) at the earth’s surface shows an increasing trend due to an accelerating depletion of stratospheric ozone. The increasing UVBR now is an important adverse environmental stressor for higher plants in semi-arid and arid lands. The photosynthetic apparatus is one of the primary target sites for UVBR. UVBR can reduce photochemical activity in photosystem II (PSII) by damaging the photosynthetic apparatus at multiple sites, including the D1 and D2 proteins.
In order to compare the responses of PSII activity between samara and leaf to enhanced UVBR, and elucidate the mechanism involved in the effect of enhanced UVBR on PSII in samara and leaf, researcher from Xinjiang Institute of Ecology and Geography and Institute of Geochemistry, Chinese Academy of Sciences, studied the effect of enhanced UVBR on PSII activity in white elm samara and leaf by measuring in vivo fast Chl a fluorescence rise transient and JIP-test.
The results show that PSII activities in both samara and leaf of white elm (U. pumila L.) were significantly inhibited by enhanced UVBR. UVBR disturbed both the donor and acceptor sides of PSII. The plastoquinone (PQ) pool size on the acceptor side, the trapped excited energy for complete reduction of QA, and the proportion of closed PSII reaction centers (RCs) increased, with PSII RCs being transformed into dissipative sinks for excitation energy under UVBR. However, samara and leaf responded to UVBR in different ways. A decrease in the F0 for leaf induced by UVBR suggests the formation of fluorescence-quenching centers. An increase in the VI for leaf under UVBR might mean the accumulation of reduced QA and PQ. F0 and VI for samara showed opposite change pattern. Leaf has the mechanism of regulation of the amount of light reaching the RC through decreasing the number of light-harvesting chlorophyll molecules under UVBR while samara may be unable to regulate the light-harvesting capacity. PSII in samara was more susceptible to UVBR than that in leaf, with PIABS for samara decreasing more rapidly by a factor of 6.4 than that for leaf. Samara can recover more easily from UVBR-induced damage to PSII than the leaf.
The research findings have been published in Russian Journal of Plant Physiology (2011, 58(5): 864–870). The article can be linked as follows: http://www.springerlink.com/content/3h1w1846880l47n6/.
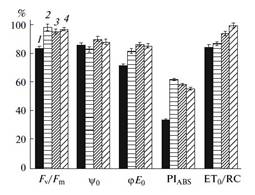
Change of PSII functional parameters for samara and leaf after 30-min UVBR and 2-h recovery after cessation of UVBR. (1) Samara after 30-min UV_B radiation; (2) samara after 2-h recovery; (3) leaf after 30-min UVBR; (4) leaf after 2-h recovery. All the values are expressed as the percentage of the control.